Водно-электролитный обмен и его нарушения : руководство для врачей / В. Г. Антонов, С. Н. Жерегеля, А. И. Карпищенко, Л. В. Минаева ; под ред. А. И. Карпищенко. - Москва : ГЭОТАР-Медиа, 2022. - 208 с. : ил. - 208 с. - ISBN 978-5-9704-6586-8. |
Аннотация
В руководстве рассмотрены вопросы обмена воды и электролитов в организме человека и их нарушения. Дан краткий обзор гомеостаза воды и электролитов в норме. Описаны варианты нарушений водно-электролитного обмена, механизмы возникновения различных нарушений, методы диагностики. Приведены значения показателей водно-электролитного обмена в норме и при патологических процессах.
Издание предназначено врачам широкого профиля, анестезиологам, реаниматологам, специалистам клинической лабораторной диагностики, студентам медицинских вузов. Книга может использоваться в системе последипломного образования врачей.
Гомеостаз натрия
В организме человека массой 70 кг содержится около 100 г натрия [7] (60 ммоль/кг). При этом 40-45% общего его количества находится в костной ткани, менее 7% приходится на внутриклеточный объем, содержание во внеклеточном пространстве составляет около 50% общего количества в организме. Активно обменивается 70% натрия. Причем обмениваемый пул включает весь натрий внутриклеточной и внеклеточной жидкостей, а также часть натрия костной ткани. Баланс натрия в организме складывается из равновесия между его поступлением, участием в биохимических и физиологических процессах, депонированием и выведением. Сбалансированность натриевого обмена оценивают по его концентрации в плазме (сыворотке) крови, моче и эритроцитах. При диареях различного генеза для более объективной оценки натриевого баланса в организме необходимо определять его потери с калом.
Поступление ионов натрия
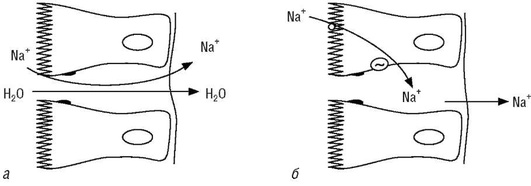
Принятый с пищей натрий легко всасывается в желудочно-кишечном тракте. Процесс всасывания начинается в желудке, однако основное количество натрия всасывается в тонкой кишке. Из гипотонических растворов натрий всасывается быстрее. Всасывание натрия осуществляется парацеллюлярно - пассивной диффузией и трансцел-люлярно - активным транспортом, который является определяющим в обмене как натрия, так и других электролитов (рис. 22).
Параллельно процессу всасывания парацеллюлярно происходит выделение натрия в просвет кишечника из крови пассивной диффузией, а также со слюной и кишечными соками. Секретируемый тем или иным путем ион натрия повторно всасывается в кровь. Выведение натрия через кишечник при сбалансированном метаболизме не превышает 2-3% его общего количества, удаляемого из организма.
Распределение ионов натрия в организме
Средние значения содержания натрия в некоторых тканях тела человека представлены в табл. 6.
| Ткань | Содержание натрия, ммоль/кг массы ткани |
|---|---|
Кости |
78 |
Зубы |
208 |
Мышцы |
31 |
Миокард |
80 |
Легкие |
108 |
Мозг |
74 |
Печень |
83 |
Почки |
76 |
Референтные интервалы
Референтные интервалы для натрия в сыворотке крови составляют от 136 до 145 ммоль/л как у детей, так и у взрослых. У недоношенных детей через 2 сут после рождения содержание натрия в крови составляет от 128 до 148 ммоль/л; в пуповинной крови у доношенных новорожденных оно равно 127 ммоль/л. Окисление компонентов смешанной пищи с освобождением энергии, эквивалентной 3000 ккал, сопровождается выделением с мочой 130-260 ммоль/л натрия. Количество натрия, удаляемого с мочой в ночное время, не превышает 20% его суточного выведения. Концентрация натрия в цереброспинальной жидкости колеблется от 136 до 150 ммоль/л, в поте - от 10 до 40 ммоль/л. Выведение натрия с калом зависит от состава продуктов питания и в среднем составляет 7,8-10,0 ммоль/сут.
Натрий выполняет следующие физиологические функции:
-
предопределяет направленный транспорт воды, электролитов, питательных веществ, продуктов обмена через эпителиальные клетки различных органов, участвуя таким образом в обмене между внешней и внутренней средами, между внутренней средой и клетками;
-
непосредственно участвует в образовании потенциала действия в нервных и мышечных клетках;
-
является основным компонентом осмотически активных частиц внеклеточной жидкости. Во внеклеточной жидкости на натриевые соли (особенно на хлорид и гидрокарбонат) приходится 90-95% общей концентрации осмотически активных частиц. При ряде заболеваний (сахарный диабет, уремия и др.) вклад натрия в концентрацию осмотически активных частиц внеклеточной жидкости снижается вследствие увеличения концентрации глюкозы, мочевины и других осмотически активных частиц;
-
определяет реальный объем внеклеточного пространства (при бессолевых диетах или при применении салуретиков внеклеточное пространство уменьшается, а при повышенном введении натрия увеличивается). Объем внутриклеточного пространства также регулируется посредством содержания натрия в плазме. При возрастании внеклеточной осмолярности (при введении гипертонических растворов NaCl) вода выводится из клеток, при потере солей клетки насыщаются водой.
Выведение ионов натрия
При нормальном потреблении пищи регуляция баланса натрия зависит от изменения скорости его экскреции почками. Натрий свободно фильтруется в почечном клубочке и реабсорбируется в различных отделах почечного канальца с участием механизмов активного и пассивного транспорта.
Количество экскретируемого натрия [E(Na+ )] определяется как разность между количеством натрия, профильтрованного в клубочках, и содержанием натрия, реабсорбированного в канальцах [R(Na+ )]. Количество профильтрованного в почечных клубочках натрия есть величина, равная произведению скорости клубочковой фильтрации (СКФ) на концентрацию его в плазме Р[(Na+ )]:
Ε(Νa+ ) = СКФ × [Р(Na+ )] - [R(Na+ )].
Теоретически результирующая величина может меняться при изменении любого из трех компонентов правой части уравнения. Однако на практике концентрация натрия в плазме крови [Р(Na+ )] мало изменяется даже при значительном нарушении натриевого баланса вследствие сопутствующего дисбаланса воды в результате ее притока из внутриклеточной жидкости или, наоборот, оттока во внутриклеточное пространство. В этой связи скорости клубочковой фильтрации и канальцевой реабсорбции выступают в качестве определяющих параметров почечного баланса натрия.
Среднесуточная скорость клубочковой фильтрации в норме составляет 120-125 мл/мин. Непрерывно в почечных клубочках фильтруется 1/5 объема почечного плазмотока, за сутки образуется до 170-180 л фильтрата. Скорость клубочковой фильтрации в дневное время, а также после приема пищи превышает среднесуточную на 30%. Напротив, в ночное время скорость клубочковой фильтрации ниже среднесуточной на 30%. Если бы реабсорбция натрия оставалась постоянной и независимой от скорости клубочковой фильтрации, то потери натрия в ночное время отсутствовали бы, а в дневное время превысили бы общее количество натрия в организме в 2 раза. Однако процессам реабсорбции натрия в канальцевой системе почки присуща гибкость, позволяющая им быстро и адекватно реагировать на изменения скорости клубочковой фильтрации, что обеспечивает сохранение соотношения между количеством профильтрованного и реабсорбированного натрия.
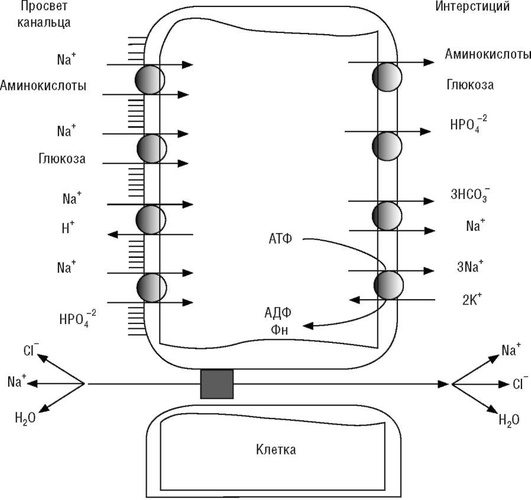
Соли натрия в клубочковом фильтрате составляют 4/5 всего количества растворенных веществ. 65% фильтруемого в клубочках натрия (натриевой нагрузки) реабсорбируется в проксимальном извитом канальце в виде NaCl (70%) и NaHCO3 (30%). Натрий удаляется из канальцевой жидкости в интерстиций через межклеточные шунты облегченной диффузией и активным транспортом. Активный транспорт натрия создает условия для его облегченной диффузии. Структура функционально единого процесса складывается из сопряженных механизмов. На люминальной мембране функционируют два механизма переноса натрия:
-
котранспорт (симпорт) - активный сопряженный транспорт через апикальную мембрану нефроцитов проксимального канальца ионов натрия в сочетании с неорганическими анионами (фосфат, сульфат, хлорид) и органическими соединениями (аминокислоты, глюкоза и др.) в цитозоль (рис. 23);
-
противоточный транспорт (антипорт) - активный сопряженный транспорт через апикальную мембрану нефроцитов проксимального канальца иона натрия в цитозоль иона водорода в канальцевую жидкость (рис. 24).
Поступление водорода в канальцевую жидкость инициирует механизм реабсорбции бикарбоната, который в итоге поступает в интерстициальное пространство. Сопутствующая трансцеллюлярная реабсорция воды предопределяет более высокую концентрацию ионов хлора в канальцевой жидкости. Ионы хлора поступают в интерстициальное пространство через межклеточные промежутки по концентрационному градиенту. В интерстициальном пространстве бикарбонат и ионы хлора предопределяют избыточный отрицательный потенциал относительно канальцевой жидкости, что способствует поступлению катионов, включая натрий, из канальцевой жидкости в интерстициальное пространство через межклеточные промежутки.
Из клетки натрий поступает в интерстициальное пространство благодаря активности Na+ ,К+ -АТФазы, которая за счет энергии АТФ обменивает три внутриклеточных иона натрия на два иона калия из интерстициального пространства, и котранспортера, который вместе с тремя ионами бикарбоната удаляет один ион натрия. Активность Na+ ,К+ -АТФазы является ключевой в функционировании ионных транспортных потоков различного вида, при этом происходит формирование электрохимического и концентрационного градиентов, использующихся различными транспортерами. Бикарбонат поступает из канальцевой жидкости в клетку посредством сопряжения с натриевыми потоками через Na+ ,Н+ -антипорт.
Вместе с бикарбонатом и натрием из канальцевой жидкости реабсорбируется вода. В канальцевой жидкости увеличивается концентрация хлора и возникает его концентрационный градиент относительно интерстиция. Хлор перемещается в интерстиций через межклеточные шунты по концентрационному градиенту, вследствие чего возникают диффузионный потенциал и электрическое поле, в котором в числе прочих катионов вслед за хлором диффундирует натрий (см. рис. 23, 24).
Взаимосвязь активного и пассивного транспорта натрия позволяет прямо и опосредованно использовать энергию гидролиза одной молекулы АТФ для реабсорбции девяти молекул натрия. Характерной особенностью процесса реабсорбции натрия (равно как и процесса реабсорбции других веществ) в проксимальном извитом канальце является то, что количество натрия, реабсорбируемого в единицу времени, не является постоянным, а составляет лишь определенный процент от его количества в фильтрате (для хлорида натрия примерно 65%), что обозначают как клубочково-канальцевое равновесие. Натрий, вследствие клубочково-канальцевого равновесия, не извлекается полностью из канальцевой жидкости при скорости клубочковой фильтрации значительно ниже оптимального физиологического уровня и, наоборот, не выводится в количествах, превышающих его содержание в организме, при скорости клубочковой фильтрации, превышающей оптимальный физиологический уровень.
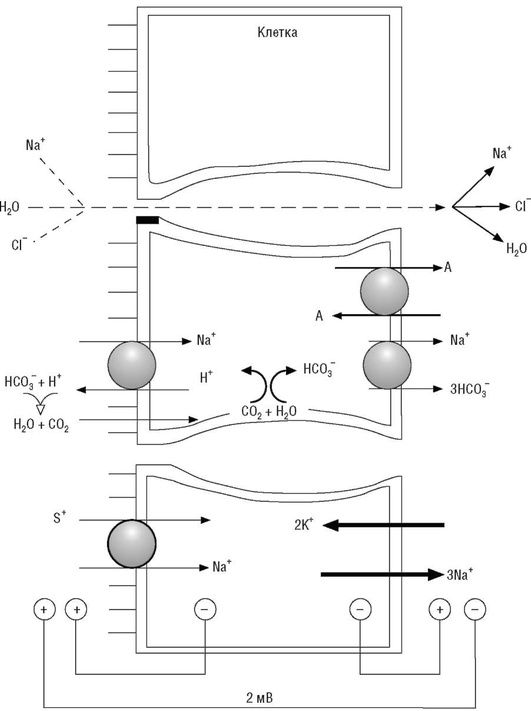
В нисходящей части петли Генле натрий не реабсорбируется, его реабсорбция возобновляется в восходящем колене по механизму котранспорта. Функциональная активность Na+ , K+ , 2Cl- -котранспортера обеспечивает реабсорбцию примерно 35% профильтровавшегося в клубочках натрия. Натрий поступает в цитозоль нефроцитов, откуда удаляется в почечный интерстиций благодаря активности Na+ ,К+ -АТФазы базолатеральной мембраны (три внутриклеточных иона натрия обмениваются на два иона калия из интерстициального пространства). Часть ионов натрия поступает в интерстиций парацеллюлярно, чему способствует преобладающий положительный заряд канальцевой жидкости относительно интерстициальной. Причиной такого распределения зарядов в канальцевой и интерстициальной жидкостях в толстом сегменте восходящего колена петли Генле является более высокая проницаемость апикальной мембраны для ионов калия в сравнении с базолатеральной мембраной, что и предопределяет формирование избыточного положительного заряда канальцевой жидкости относительно интерстициальной жидкости. В этой части канальца (как и в проксимальном извитом канальце) Na+ ,К+ -АТФаза освобождает энергию АТФ для реабсорбции Na+ (удаление из цитозоля трех ионов натрия создает электрохимический и концентрационный градиенты для транспорта натрия с участием Na+ , K+ , 2Cl- -котранспортера, рис. 25).
К дистальной части канальца в его жидкости остается 2% профильтровавшегося натрия. Транспорт ионов натрия в дистальном канальце и кортикальной части собирательной трубки через апикальную мембрану нефроцита в клетку происходит посредством работы симпорта - котранспорт с ионами хлора - и антипорта, осуществляющего обмен натрия канальцевой жидкости на протон водорода из цитозоля. Благодаря работе нескольких систем активного транспорта ионов натрия его удаление из канальцевой жидкости идет интенсивнее, чем удаление ионов хлора, что предопределяет формирование отрицательного заряда канальцевой жидкости относительно интерстициальной. Из цитозоля нефроцитов ион натрия удаляется в почечный интерстиций благодаря активности Na+ ,К+ -АТФазы базолатеральной мембраны (три внутриклеточных иона натрия обмениваются на два иона калия из интерстициального пространства). Вследствие работы Na+ ,К+ -АТФазы базолатеральной мембраны нефроцитов между канальцевой жидкостью и цитозолем создаются электрохимический и концентрационный градиенты, обеспечивающие транспорт натрия через люминальную мембрану по натриевым каналам через симпорт с Cl- и антипорт с Н+ (рис. 26).
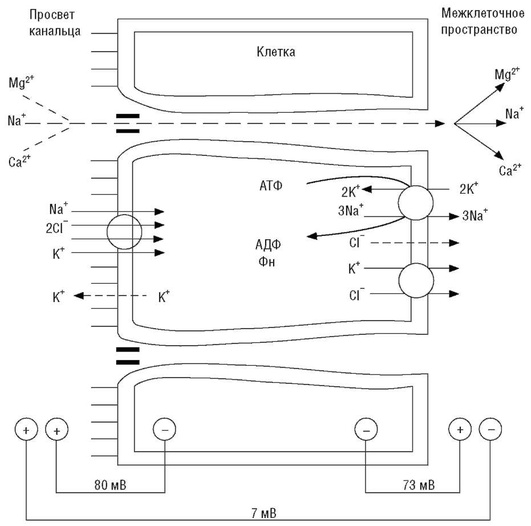
Особенностью транспортных процессов в дистальной части канальца является их доступность для регуляторных воздействий. Неравновесная реабсорбция ионов натрия и хлора усиливается при действии альдостерона, так как он стимулирует процесс реабсорбции натрия в обмен на ионы водорода. Альдостерон стимулирует синтез и встраивание в апикальную мембрану дополнительных молекул антипортера, благодаря чему экскретируемое с мочой количество натрия (в норме 50-130 ммоль/л) может как падать до значений, близких к нулю, так и кратно превышать референтные значения.
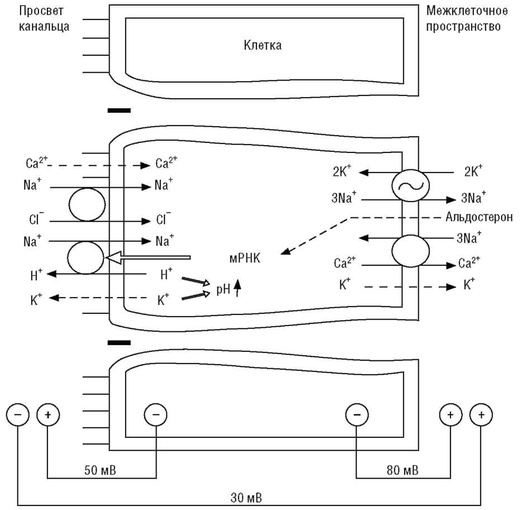
Регуляция баланса натрия
Селективная регуляция содержания натрия осуществляется нейро-гуморальными механизмами. Инициирующими сигналами дисбаланса в обмене натрия являются изменения объема жидкости и (или) концентрации натрия, калия в плазме крови, снижение реабсорбции натрия в проксимальной части канальца и восходящем колене петли Генле. Уменьшение объема внеклеточной жидкости приводит к раздражению рецепторов, реагирующих на изменения объема (в системе низкого давления - рецепторы левого предсердия, в системе высокого давления - барорецепторы каротидного синуса и дуги аорты), внутрипочечных барорецепторов, реагирующих на снижение перфузионного давления. Снижение реабсорбции натрия в проксимальной части канальца и восходящем колене петли Генле увеличивает количество натрия, достигающего плотного пятна (macula densa), и вызывает секрецию ренина. Гуморальными эффекторами реакции центральной нервной системы на сигнал с соответствующих рецепторов являются альдостерон и предсердный натрийуретический фактор (предсердный натрий-уретический пептид). Параллельно с этими регуляторными факторами функционируют другие механизмы регуляции натриевого баланса, связанные с перераспределением кровотока между корковым и мозговым веществом почек, эффектами различных системных и почечных гормонов. Итоговым результатом функционирования системных и специфических механизмов регуляции баланса натрия будет восстановление физиологического равновесия.
Альдостерон
Альдостерон является гормоном, синтезируемым клубочковой зоной коры надпочечников, относится к минералокортикоидам, осуществляет специфическую регуляцию водно-солевого гомеостаза. Установлено, что рецепторы к глюко- и минералокортикоидам имеют близкую структуру, поэтому минералокортикоидные рецепторы обладают одинаковым сродством как к альдостерону, так и к кортизолу.
Физиологическая специфичность действия альдостерона на водно-солевой гомеостаз обеспечивается тем, что в клетках-мишенях при воздействии альдостерона экспрессируется ген, кодирующий фермент 11β-гидроксистероиддегидрогеназу второго типа (HSD 11β2). Этот фермент трансформирует кортизол в кортизон, не имеющий сродства к минералокортикоидным рецепторам. В плазме крови не обнаружено специфического белка-переносчика, что, вероятно, обусловливает быстрое удаление альдостерона из крови клетками печени. Продукция альдостерона клубочковой зоной зависит от:
-
концентрации натрия в плазме (секреция альдостерона увеличивается в ответ на снижение уровня натрия в плазме);
-
концентрации калия в плазме (увеличение концентрации калия в плазме стимулирует секрецию альдостерона; напротив, гипокалиемия ее снижает);
-
интенсивности образования ангиотензина II (образование ангиотензина II стимулируется ренином, образующимся в результате снижения почечного перфузионного давления; ангиотензин II среди прочих эффектов приводит к повышению интенсивности синтеза альдостерона);
-
секреции АКТГ (АКТГ обладает пермиссионным эффектом на секрецию альдостерона).
Клетки-мишени альдостерона выявлены в дистальных извитых канальцах и собирательных трубках. Связывание гормона осуществляется белками - рецепторами цитоплазмы. Конкретный молекулярный механизм действия альдостерона не вполне ясен, однако очевидно, что его присутствие стимулирует активность Na+ ,Κ+ -АТФазы, ряда мито-хондриальных ферментов и скорость оборота АТФ, увеличивает количество и проницаемость натриевых каналов на люминальной мембране.
Совокупность всех этих эффектов альдостерона в норме приводит к активной адсорбции и транспорту ионов натрия из просвета канальцев в интерстициальную жидкость в обмен на ионы калия и водорода.
Предсердный натрийуретический пептид
Название предсердный натрийуретический пептид (ANP) объединяет группу пептидов, обладающих выраженной натрийуретической активностью и образующихся из общего пептида-предшественника - препроANP, состоящего из 151 аминокислоты. ANP включает кардионатрины, атриопептины или аурикулины. Его синтез в основном осуществляется в клетках предсердий, небольшое количество образуется в клетках желудочков. Некоторые количества ANP синтезируются в легких, а также нейронами центральной и периферической нервной системы. В плазме крови ANP циркулирует в форме пептида из 28 аминокислот, 17 из которых формируют кольцо через дисульфидный мостик. ANP секретируется в ответ на растяжение предсердий (увеличение объема внутрисосудистой жидкости при различных патологических состояниях, изменение положения тела из вертикального в горизонтальное, физическая нагрузка). Выработка ANP возрастает под влиянием глюкокортикоидов, вазопрессина, эндотелина или α-адренорецепторов. Рецепторы к ANPА и ANPВ связаны с гуанилатциклазой, а рецепторы ANPС ответственны за элиминацию пептидов. ANP подавляет секрецию ренина, альдостерона и вазопрессина, увеличивает скорость клубочковой фильтрации и снижает реабсорбцию натрия и воды.
Период полувыведения ANP из крови составляет 3-5 мин, он связывается с рецептором ANPС и в цитозоле тканей разрушается нейтральной эндопептидазой.
Другие механизмы регуляции баланса натрия
Количество выделяемого из организма натрия может регулироваться перераспределением кровотока между корковым и мозговым веществом почки. Особенностью функциональной активности «кортикального» нефрона является большая способность к выделению натрия, напротив, глубокий юкстамедуллярный нефрон способен реабсорбировать большие количества натрия. Перераспределение кровотока в корковую или медуллярную часть почечной ткани будет приводить к натрийурезу или задержке натрия.
К снижению выведения натрия приводит возбуждение симпатической нервной системы, волокна которой иннервируют проксимальную часть канальца. Подобным образом действуют и глюко-кортикоиды, гормон роста и инсулин. Напротив, паратиреоидный гормон, прогестерон, глюкагон, простагландины стимулируют натрийурез. Молекулярные основы натрий-сберегающего (равно как и натрийуретического) действия перечисленных гормонов не вполне ясны. Механизм регуляторного действия ряда из них может быть связан с расширением или, напротив, сужением перитубулярных капилляров, следствием этого будет изменение соотношения между гидростатическим и онкотическим давлениями, что в итоге может либо способствовать реабсорции натрия, либо стимулировать натрийурез.
Нарушения баланса натрия
Нарушения физиологически оптимального натриевого обмена могут привести к состоянию его избыточного или недостаточного количества в организме. Натриевый баланс организма оценивается по концентрации натрия в крови. Если количество натрия в крови превышает референтное значение, то говорят о гипернатриемии, и напротив, если количество натрия ниже его, то имеет место гипонатриемия.
Гипернатриемия
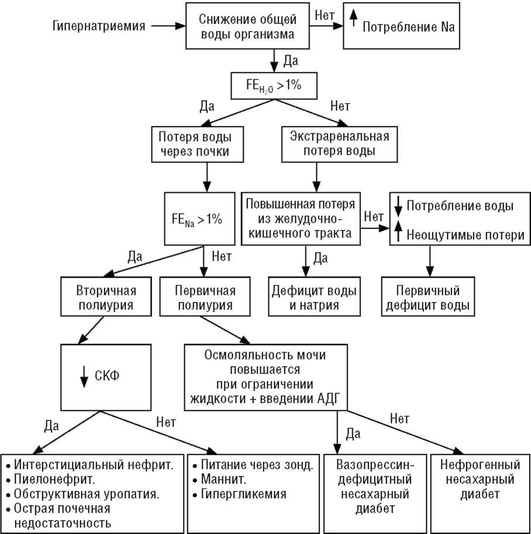
Состояние гипернатриемии характеризуется устойчивым повышением концентрации натрия в крови более 150 ммоль/л. Оно может быть следствием как избыточного количества натрия в крови при нормальном объеме воды, так и недостаточного объема воды относительно нормального количества натрия. Гипернатриемия независимо от механизма ее развития приводит к уменьшению объема внутриклеточного пространства. Дисбаланс между физиологически оптимальным соотношением воды и натрия может быть следствием двух причин:
Расстройства водного баланса при гипернатриемии аналогичны гипертонической дегидратации.
Гипернатриемия как следствие избыточного введения натрия и (или) недостаточного поступления воды
Избыточное введение натрия при относительно меньших объемах потребляемой жидкости индуцирует секрецию гуморальных факторов, действие которых направлено на сохранение осмотически свободной воды и удаление избытка натрия. При максимальной концентрирующей способности почек потери облигатной воды являются неизбежными. Если не будет восстановлен прием воды, пропорциональный количеству натрия, развивается состояние гипернатриемии. Оно развивается тем быстрее, чем выше потери воды.
Гипернатриемия как следствие потери воды
Различают почечные и непочечные потери воды. Почечные потери воды могут быть связаны с потерей облигатной воды - вторичная полиурия и (или) осмотически свободной воды - первичная полиурия. Преимущественная потеря воды (но не натрия) может быть обусловлена внепочечными механизмами, в частности через желудочно-кишечный тракт.
Потеря облигатной воды (вторичная полиурия). Факторами, нарушающими реабсорбцию воды в проксимальном канальце и нисходящем колене петли Генле, являются низкомолекулярные мало- и нереабсорбируемые соединения. Они предупреждают всасывание воды из канальцевой жидкости, создавая в ней осмотическую силу, противодействующую осмотической силе реабсорбируемого натрия. Подобное явление называют осмотическим диурезом, а вещества - осмотическими диуретиками. В качестве примера можно привести осмотический диуретик маннитол (Маннит♠ ), который хорошо фильтруется в клубочке и практически не реабсорбируется в канальце. В проксимальной части канальца реабсорбция натрия обеспечивает изоосмотическое всасывание воды и уменьшение объема канальцевой жидкости. Уменьшение объема канальцевой жидкости приведет к увеличению концентрации Маннита♠ до точки, когда создаваемая им осмотическая сила превысит осмотическую силу реабсорбируемого натрия и, соответственно, остановит изоосмотическую реабсорбцию воды. В результате этого в проксимальном канальце часть воды канальцевой жидкости в присутствии Маннита♠ не реабсорбируется. Реабсорбция воды снижается и в нисходящей части петли Генле.
Механизм нарушения противоточного всасывания воды обусловлен как наличием противодействующей осмотической силы канальцевой жидкости, так и снижением осмолярности медуллярного интерстиция. Последний эффект нереабсорбируемые соединения создают опосредованно. Не реабсорбированная в проксимальном канальце вода увеличивает объем канальцевой жидкости и уменьшает концентрацию натрия. В свою очередь это уменьшает концентрационный градиент натрия относительно внутриклеточного пространства и снижает активность Na+ , K+ , 2Cl- -котранспортера. Итоговым результатом является снижение осмоляльности медуллярного интерстиция и соответственно уменьшение количества реабсорбируемой воды. Не реабсорбированная в проксимальном канальце и нисходящей части петли Генле вода выводится из организма. Механизм снижения реабсорбции воды при действии осмотических диуретиков сопряжен с потерей натрия. Однако потери воды более значительны, вследствие чего осмотический диурез приводит к гипернатриемии. Подобно Манниту♠ осмотический диурез вызывают декстроза (Глюкоза♠ ) и мочевина при концентрациях, превышающих пороговые величины реабсорбции.
Потеря осмотически свободной воды почками (первичная полиурия).
Она может быть связана как с поражением дистальной части канальца и кортикального отдела собирательной трубки, так и с недостаточной секрецией и (или) снижением чувствительности рецепторов канальцевого аппарата к антидиуретическому гормону. Независимо от причины удаление осмотически свободной воды почками приводит к увеличению внеклеточной концентрации натрия и состоянию гипернатриемии.
Потеря воды через желудочно-кишечный тракт. Концентрация натрия в желудочном секрете значительно ниже его концентрации в плазме крови (см. табл. 5). Потеря желудочного секрета при рвоте или фистуле желудка сопровождается относительно меньшим удалением из организма натрия, чем воды. Интестинальные секреты в норме сравнимы по содержанию натрия с плазмой крови, однако при заболеваниях (поражениях) того или иного отдела кишечника количество натрия в них может значительно снижаться. Следствием потери интестинальных секретов, обедненных натрием, является развитие гипернатриемии.
Лабораторные данные при гипернатриемии
При гипернатриемии концентрация натрия в плазме крови более 150 ммоль/л сочетается с повышенной осмоляльностью плазмы. Концентрация натрия в моче ниже 10 ммоль/л, осмоляльность мочи выше осмоляльности плазмы. При почечных потерях суточная экскреция воды превысит 1% количества фильтруемой жидкости (полиурия), напротив, при внепочечных потерях ее относительное содержание в экскретируемой фракции будет в пределах референтных значений. Количество экскретируемого натрия при вторичной полиурии превысит 1% его количества в фильтруемой фракции, при первичной полиурии, напротив, относительное содержание натрия в экскретируемой фракции будет в пределах референтных значений. Дегидратационный тест (введение АДГ при ограничении приема жидкости: осмоляльность мочи увеличивается - тест положительный; осмоляльность мочи не меняется - тест отрицательный) при первичной полиурии, обусловленной дефицитом АДГ, положительный, напротив, при поражениях дистальных отделов канальца дегидратационный тест отрицательный. Возможный диагностический алгоритм варианта гипернатриемии представлен на рис. 27.
Гипонатриемия
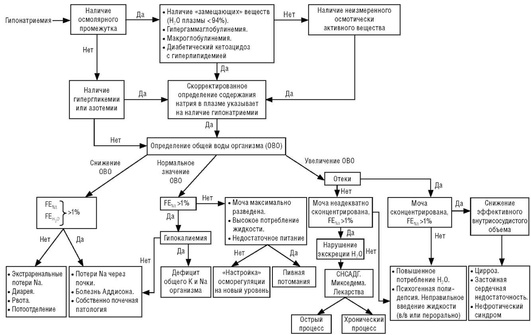
Состояние гипонатриемии характеризуется устойчивым понижением концентрации натрия в крови менее 135 ммоль/л. Гипонатриемия может быть следствием как избыточного объема воды относительно нормального количества натрия в крови, так и недостаточного его количества относительно нормального объема воды. Гипонатриемия независимо от механизма ее развития приводит к увеличению объема внутриклеточного пространства, так как оно оказывается гипертоничным по отношению к интерстициальной жидкости. Гипонатриемия в сочетании с состоянием гипергидратации рассмотрена в разделе «Гомеостаз воды в организме». Здесь остановимся на других вариантах дисбаланса между физиологически оптимальным соотношением воды и натрия, которые могут быть следствием:
Псевдогипонатриемия
Характеризуется пониженной концентрацией натрия в единице объема плазмы, однако пересчет концентрации на единицу объема воды дает нормальные значения. Подобное расхождение в удельной концентрации натрия обусловлено возрастанием объема неводного белкового и (или) липидного компонента плазмы крови. В норме количество воды в единице объема плазмы составляет 94%. При сахарном диабете, гипертермии при инфекционных процессах, операционном стрессе, макроглобулинемии, множественной миеломе и ряде других заболеваний в единице объема плазмы крови увеличивается доля белкового и (или) липидного компонента, что соответственно понижает количество воды в единице объема плазмы. Последнее может быть рассчитано по следующей формуле:
Ws = 99,1 - 1,03 × Ls - 0,73 × Ps,
где W s - количество воды в 100 мл плазмы крови, мл; 99,1 - объем плазмы без кристаллических (нелипидных и небелковых) твердых частиц; Ls - концентрация общих липидов в плазме крови, мг/дл; Ps - концентрация общего белка в плазме крови, г/дл; 1,03 и 0,73 - коэффициенты.
Увеличение содержания липидов и (или) белков в 1 дл плазмы крови уменьшает количество воды в этом объеме плазмы крови и соответственно натрия, растворимого только в водной фазе. При псевдо-гипонатриемии практически не меняется измеренная осмоляльность, однако расчетная осмоляльность, в значении которой учитываются только водорастворимые фракции, снижается. В случае гипергликемии значения расчетной и измеренной осмоляльности будут соответствовать нормальным, что обусловлено осмотической активностью молекул глюкозы, в этом случае будет отмечаться истинная гипонатриемия.
Преобладающие потери натрия относительно потерь воды
Эти изменения отмечаются при интестинальных потерях жидкости, когда, например, количество натрия в секрете тощей и подвздошной кишки превышает количество натрия в плазме (см. табл. 5). Если происходит селективная потеря секретов из этих отделов кишки, то потери натрия более значительны, чем потери воды, и в результате этого развивается состояние гипонатриемии.
Гипонатриемия и нормальный объем внеклеточной воды
Гипонатриемия при нормальном объеме внеклеточной жидкости может быть обусловлена избирательным нарушением функции почек по сохранению натрия. Водовыделительная функция почек при этом остается в норме. При нормальной функции почек избыточное выделение натрия и сохранение воды имеют место при онкологических заболеваниях, туберкулезе, алкоголизме. Полагают, что организм при этих заболеваниях настраивается на новый уровень осморегуляции. Предположение обосновывается тем, что при туберкулезе и онкологических заболеваниях меняется спектр внутриклеточных осмотически активных соединений, уменьшается концентрация калия. Снижение общего количества калия в организме приводит к гипонатриемии. Механизм регуляторного взаимодействия между содержанием внутриклеточного калия и внеклеточного натрия неизвестен. Водовыделительная функция почек при дефиците калия не страдает, в то время как чувствительность к регуляторным влияниям антидиуретического гормона повышена.
При нормальной функции почек к гипонатриемии приводит употребление больших количеств обедненной натрием жидкости. В качестве примера в литературе обсуждается гипонатриемия в результате ежедневного приема 5 л и более пива. Механизм развития гипонатрие-мии в этой ситуации обусловлен тем, что достижение нижнего предела разведения мочи при избыточном поступлении гипоосмоляльной жидкости достигается выведением натрия, что с течением времени приводит к гипонатриемии.
Лабораторные данные при гипонатриемии
Измерение и расчет осмолярности, определение отношения концентрации натрия в плазме к измеренной в ней осмолярности ([Na+ ]/[Pосм ]) и осмотического промежутка необходимы для диагностики псевдо-гипонатриемии. С учетом основного вклада натрия в значение расчетной осмолярности отмечается ее снижение при псевдонатриемии. Осмотическая активность избыточных количеств белка и липидов практически мало сказывается на изменении измеряемой осмолярности. Сочетание осмотических параметров плазмы крови при псевдогипонатриемии характеризуется увеличением осмотического промежутка. Значение [Na+ ]/[Pосм ] уменьшается. В норме значение этого показателя варьирует в пределах 0,43-0,50 ед. Низкое содержание натрия в плазме крови и падение величины [Na+ ]/[Pосм ] к измеренной осмолярности ниже 0,43 наблюдаются преимущественно при гиперлипемии и гиперпротеинемии. Такое снижение отношения [Na+ ]/[Pосм ] при нормальной концентрации натрия и глюкозы указывает на накопление преимущественно азотистых метаболитов в крови.
При гипонатриемии, связанной с непочечными потерями воды и натрия, в крови отмечается выраженное увеличение азота мочевины на фоне нормальных значений креатинина. Суточный диурез снижается при концентрации натрия в моче менее 10 ммоль/л. Экскретируемые фракции натрия и воды составляют менее 1% фильтруемого количества.
Гипонатриемия при нормальном объеме внеклеточного пространства характеризуется снижением концентрации натрия и калия в плазме крови. Прием натрия при этом виде расстройств натриевого баланса приводит к натрийурезу.
Возможный диагностический алгоритм варианта гипонатриемии с учетом состояния гипонатриемии при гипергидратации (см. раздел «Гомеостаз воды в организме») представлен на рис. 28.
Измеряемая концентрация натрия в плазме крови при гипонатриемии позволяет рассчитать его общий дефицит и количество, необходимое для его восполнения.
Расчет дефицита натрия плазмы, ммоль/л:
Деф. Na+ ~ (142 - [Na+ пл ]) × МТ × 0,2,
где [Na+ пл ] - измеряемая (действительная) концентрация натрия в плазме крови, ммоль/л; МТ - масса тела, кг.
Пример: [Na+ пл ] - 120 ммоль/л, МТ - 70 кг.
Деф. Na+ ~ (142 - 120) × 70 × 0,2 ~ 308 ммоль/л.
Полученный результат указывает на общий дефицит Na+ около 308 ммоль/л. Количество Na+ легко перевести в единицы массы, умножив на 23 (молекулярная масса 1 ммоля натрия составляет 23 мг).
Na+ ~ 308 × 23 ~ 7084 мг или ~ 7 г.
Такое количество Na+ содержится в 18 г натрия хлорида, так как именно он используется для терапии.
Расчет необходимого объема 10% натрия хлорида, мл, для коррекции гипонатриемии проводится по формуле:
V(10%NaCl) ~ (142 - [Na+ пл ]) × МТ × 0,12. Пример: [Na+ пл ] - 120 ммоль/л, МТ - 70 кг.
V(10%NaCl) ~ (142 - 120) × 70 × 0,12 ~ 185 мл. 185 мл 10% раствора натрия хлорида содержат ~18 г натрия хлорида и соответственно ~7 г Na+ .